简答题
正确答案
半乳糖操作子。
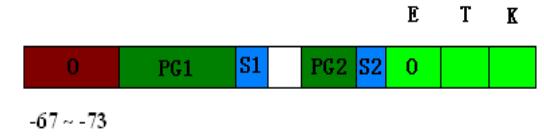
包括三个结构基因:异构酶(galE),半乳糖-磷酸尿嘧啶核苷转移酶(galT),半乳糖激酶(galK)。这3个酶的作用使半乳糖变成葡糖-1-磷酸。半乳糖操作子有两个启动子,PG1和PG2。两个RNA聚合酶结合位点S1和 S2(转录起始点),mRNA可以从不同的起始点开始转录。它有两个O区,一个在P区上游-67~-73,而不是在P区与结构基因之间,另一个O区在结构基因galE内部。
作用机制:从S1起始的转录只有当培养基中无葡萄糖时才能进行。从 S2起始的转录要依赖于葡萄糖,高水平的CAP能够抑制从S2起始的转录。当有CAP时,转录从从S1起始,无CAP时,转录从S2起始。半乳糖对细菌有双重作用,一方面可以作为碳源供细胞生长,另一方面又是细胞壁的成分。所以需要一个不依赖于CAP的启动子(S2 )进行本底水平的永久型合成。同时以半乳糖作为碳源供细胞生长,需要一个依赖CAP的启动子(S1)对高水平合成进行调节。
色氨酸操纵子。
合成色氨酸所需酶类的基因E、D、C、B、A,受其上游调控蛋白R的调控。R并没有与O结合的活性,只有当环境能提供足够浓度的色氨酸时,R与色氨酸结合后而活化,能够与O结合,阻遏结构基因的转录,操纵子由开到关。
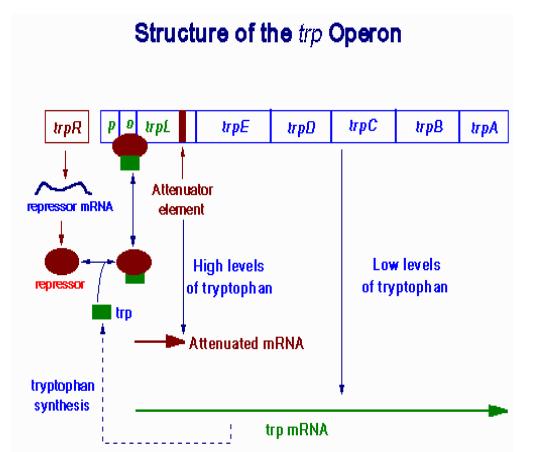
当色氨酸达到一定浓度,但还没有高到能够活化R使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,而且产生的酶量与色氨酸浓度呈负相关。这种调控现象与色氨酸操纵子特殊的结构有关。O与结构基因trpE之间有140bp的一段先导序列。当Trp浓度升高,能够阻止RNA聚合酶的转录。先导序列含有3对反向重复序列,在被转录生成mRNA时都能够形成发夹式结构,使转录终止。
阻遏步骤:
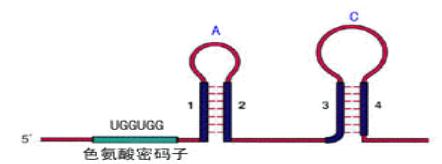
A. 当色氨酸浓度低时,生成的tRNAtrp量就少,使核糖体沿mRNA翻译移动的速度慢,赶不上RNA聚合酶沿DNA移动转录的速度,这时核糖体占据1位的机会较多,使1、2不能配对,2、3配对,阻止了3、4生成终止信号的结构,trp操纵元处于开放状态。
B. 当色氨酸浓度增高时,核糖体沿mRNA翻译移动的速度加快,占据到2段的机会增加,2、3配对的机会减少,3、4形成终止结构的机会增多,转录减弱。
C. 当所有氨基酸都不足时,核糖体翻译移动的速度就更慢,甚至不能占据1的序列,结果有利于1、2和3、4发夹结构的形成,于是转录停止。
阿拉伯糖操纵子。
阿拉伯糖操的降解需要三个基因:araB、araA、araD。形成一个基因簇araBAD。它们分别编码三个酶:核酮糖激酶,L-阿拉伯糖异构酶,L-核酮糖-5-磷酸-4-差向异构酶。在这个操纵子中阿拉伯糖的代谢顺序是以araA,araB,araD的序列进行。与araBAD相邻的是一个复合的启动子区域、两个操纵区(O1、O2)和一个调节基因araC。AraC蛋白同时显示正、负调节因子的功能。
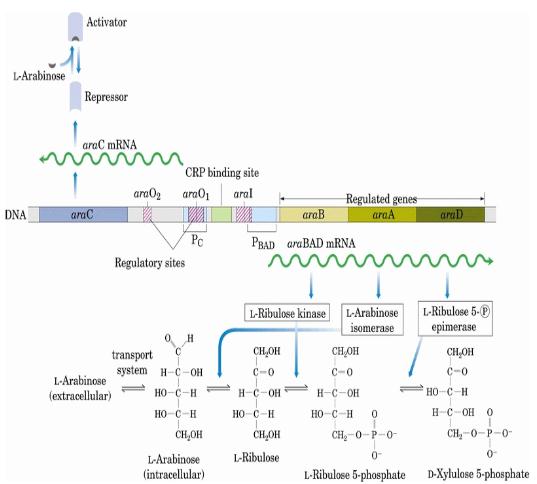
调控机制:a、当细胞内没有araC蛋白时,由PC启动子起始araC基因转录。
b、当体系中葡萄糖的水平较高时,阿拉伯糖水平较低时,AraC蛋白与操纵区O2以及以及araI诱导因子结合区上半区相结合,形成DNA回转结构,araBAD基因不表达。
c、当体系中有阿拉伯糖但无葡萄糖存在时,AraC蛋白与阿拉伯糖相结合,改变构象成为激活蛋白,AraC蛋白同源二聚体分别与ara O1和araI结合,DNA回转结构被破坏,RNA聚合酶在AraC蛋白和CRP-cAMP的共同作用下起始PBAD所调控的结构基因的表达。
包括三个结构基因:异构酶(galE),半乳糖-磷酸尿嘧啶核苷转移酶(galT),半乳糖激酶(galK)。这3个酶的作用使半乳糖变成葡糖-1-磷酸。半乳糖操作子有两个启动子,PG1和PG2。两个RNA聚合酶结合位点S1和 S2(转录起始点),mRNA可以从不同的起始点开始转录。它有两个O区,一个在P区上游-67~-73,而不是在P区与结构基因之间,另一个O区在结构基因galE内部。
作用机制:从S1起始的转录只有当培养基中无葡萄糖时才能进行。从 S2起始的转录要依赖于葡萄糖,高水平的CAP能够抑制从S2起始的转录。当有CAP时,转录从从S1起始,无CAP时,转录从S2起始。半乳糖对细菌有双重作用,一方面可以作为碳源供细胞生长,另一方面又是细胞壁的成分。所以需要一个不依赖于CAP的启动子(S2 )进行本底水平的永久型合成。同时以半乳糖作为碳源供细胞生长,需要一个依赖CAP的启动子(S1)对高水平合成进行调节。

色氨酸操纵子。
合成色氨酸所需酶类的基因E、D、C、B、A,受其上游调控蛋白R的调控。R并没有与O结合的活性,只有当环境能提供足够浓度的色氨酸时,R与色氨酸结合后而活化,能够与O结合,阻遏结构基因的转录,操纵子由开到关。

当色氨酸达到一定浓度,但还没有高到能够活化R使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,而且产生的酶量与色氨酸浓度呈负相关。这种调控现象与色氨酸操纵子特殊的结构有关。O与结构基因trpE之间有140bp的一段先导序列。当Trp浓度升高,能够阻止RNA聚合酶的转录。先导序列含有3对反向重复序列,在被转录生成mRNA时都能够形成发夹式结构,使转录终止。

阻遏步骤:
A. 当色氨酸浓度低时,生成的tRNAtrp量就少,使核糖体沿mRNA翻译移动的速度慢,赶不上RNA聚合酶沿DNA移动转录的速度,这时核糖体占据1位的机会较多,使1、2不能配对,2、3配对,阻止了3、4生成终止信号的结构,trp操纵元处于开放状态。
B. 当色氨酸浓度增高时,核糖体沿mRNA翻译移动的速度加快,占据到2段的机会增加,2、3配对的机会减少,3、4形成终止结构的机会增多,转录减弱。
C. 当所有氨基酸都不足时,核糖体翻译移动的速度就更慢,甚至不能占据1的序列,结果有利于1、2和3、4发夹结构的形成,于是转录停止。
阿拉伯糖操纵子。
阿拉伯糖操的降解需要三个基因:araB、araA、araD。形成一个基因簇araBAD。它们分别编码三个酶:核酮糖激酶,L-阿拉伯糖异构酶,L-核酮糖-5-磷酸-4-差向异构酶。在这个操纵子中阿拉伯糖的代谢顺序是以araA,araB,araD的序列进行。与araBAD相邻的是一个复合的启动子区域、两个操纵区(O1、O2)和一个调节基因araC。AraC蛋白同时显示正、负调节因子的功能。

调控机制:a、当细胞内没有araC蛋白时,由PC启动子起始araC基因转录。
b、当体系中葡萄糖的水平较高时,阿拉伯糖水平较低时,AraC蛋白与操纵区O2以及以及araI诱导因子结合区上半区相结合,形成DNA回转结构,araBAD基因不表达。
c、当体系中有阿拉伯糖但无葡萄糖存在时,AraC蛋白与阿拉伯糖相结合,改变构象成为激活蛋白,AraC蛋白同源二聚体分别与ara O1和araI结合,DNA回转结构被破坏,RNA聚合酶在AraC蛋白和CRP-cAMP的共同作用下起始PBAD所调控的结构基因的表达。
答案解析
略
相似试题
除了乳糖操纵子学说解释了原核生物基因表达调控的原理外,您是否知道解释原核生物基因表达调控的其他学说?如果知道,请作简单介绍。
简答题查看答案
除了乳糖操纵子学说解释了原核生物基因表达调控的原理外.您是否知道解释原核生物基因表达调控的其他学说?如果知道,请作简单介绍。
简答题查看答案
原核生物基因表达调控,阻遏蛋白与效应物结合时,基因不转录();阻遏蛋白不与效应物结合时基因不转录();有效应物时,激活蛋白处于无活性状态基因不转录();有效应物时,激活蛋白处于活性状态基因转录()。
填空题查看答案
原核生物中多基因组成的基因表达和调控元件称为()
单选题查看答案
原核生物中的多基因组成的基因表达和调控单元,称为()。
单选题查看答案
不同生物使用不同的信号来指挥基因调控。原核生物中()和()对基因表达起着举足轻重的影响。在高等真核生物中()和()是基因表达调控的最主要手段。
填空题查看答案
简述原核生物基因表达的调控的特点。
简答题查看答案
原核生物基因表达的调控主要发生在()。
填空题查看答案
比较原核生物和真核生物基因表达调控的异同。
简答题查看答案